
ЛОКАЛИЗАЦИЯ ПСИХИЧЕСКИХ ФУНКЦИЙ в коре головного мозга
Расстановка ударений: ЛОКАЛИЗА`ЦИЯ ПСИХИ`ЧЕСКИХ ФУ`НКЦИЙ в коре головного мозга
ЛОКАЛИЗАЦИЯ ПСИХИЧЕСКИХ ФУНКЦИЙ в коре головного мозга - связь психич. функций с отдельными областями и участками коры больших полушарий головного мозга.
В течение многих столетий философы и учёные пытались "локализовать" сложные психич. функции в определённых ограниченных участках мозга. Так, ещё в ср. века было высказано предположение, что материальным субстратом психич. "способностей" является жидкость, заполняющая полости, расположенные внутри больших полушарий мозга (т. н. мозговые желудочки), причём считали, что передний мозговой желудочек является носителем способности восприятия, средний - воображения и интеллекта, а задний - памяти. В 17 в. франц. философ Р. Декарт выдвинул предположение, что материальной основой психики является шишковидная железа, расположенная в центральной части основания полушарий мозга; по его мнению, она выделяет особые жидкости (spiritus animales), к-рые, циркулируя по нервным трубкам, являются субстратом психики. В дальнейшем в качестве мозгового субстрата психич. процессов стали рассматривать плотное вещество больших полушарий мозга, причём одни авторы были склонны видеть материальную основу психич. процессов в белом веществе, составляющем массу больших полушарий, а другие - в сером веществе, облегающем снаружи большие полушария, - в коре головного мозга. Первым, кто высказал мысль, что субстратом психич. процессов является кора головного мозга, был австр. анатом Ф. Галль (1758 - 1828); он предполагал, что различные сложные психич. "способности" размещены в изолированных участках коры головного мозга и что чем больше разрастаются эти участки, тем больше развита у человека соответствующая "способность". Эти фантастич. предположения Галля легли в основу особой созданной им "науки" - френологии, к-рая якобы позволяла по выпуклостям черепа определять "способности" человека и была иллюстрирована в наивных "функциональных картах мозга" (см. рис. 1). Френология вызвала резкую критику ряда учёных (наир., франц. физиолога М. Флуранса, 1794 - 1867), к-рые высказали обратное предположение - что мозг работает как единое целое и что в мозговой коре нельзя выделить изолированных участков, являющихся субстратом отдельных "способностей". В отличие от "узколокализационистских" взглядов Галля, эти теории получили название "антилокализационизма". Дальнейшие наблюдения над нарушением психич. процессов при поражениях отд. участков коры головного мозга и физиологич. исследования с раздражением отд. зон мозговой коры показали, однако, что кора головного мозга вовсе не является однородной массой и что её отдельные части связаны с различными функциями. Ещё в 1861 франц. анатом П. Брока указал на то, что поражение задней части 3-й лобной извилины левого полушария вызывает нарушение моторной речи (т. н. моторная афазия), и высказал предположение, что в этой области хранятся "моторные образы слов". В 1873 нем. психиатр К. Вернике показал, что поражение задней трети верхней височной извилины левого полушария приводит к нарушению возможности слышать и понимать чужую речь (т. н. сензорная афазия), и высказал предположение, что эта часть мозговой коры является субстратом "сензорных образов слов". Предположения о функциональной неоднозначности различных участков мозговой коры получили подтверждение в физиологич. и анатомич. данных. В 1871 нем. физиологи Фрич и Гитциг показали, что раздражение отд. участков передней центральной извилины мозга приводит к сокращению конечностей противоположной стороны, выделив т. о. двигательные зоны коры головного мозга. Эти предположения были в дальнейшем подтверждены англ. физиологом Ч. Шеррингтоном. Развитие нейрохирургии позволило провести подобные наблюдения на человеке. Полученные данные показали, что раздражение передней центральной извилины вызывает сокращение конечностей противоположной стороны тела, причём раздражение верхних участков этой области вызывает движения ноги, средних - руки, нижних - мускулатуры лица. Аналогичные данные были получены и при раздражении др. участков мозговой коры. Так, раздражение отд. участков задней центральной извилины вызывало ощущение "мурашек" или "тока" в соответствующих частях противоположной стороны тела, причём так же, как это имело место при раздражении передней центральной извилины, раздражение верхних участков этой области вызывало такие ощущения в противоположной ноге, средних - в противоположной части туловища и руке, нижних - в противоположной части лица. Это показало, что задняя центральная извилина является "чувствительной зоной" мозговой коры. Аналогичные наблюдения, проведённые рядом исследователей (О. Петцль, Г. Хофф, У. Пенфилд и др.), показали, что существуют и другие высокоспециализированные участки коры головного мозга. Так, раздражение коры затылочной области вызывало появление зрительных ощущений (см. рис. 2), а раздражение верхних отделов коры височной области - слуховые ощущения. Важнейшим подтверждением функциональной неоднозначности различных участков мозговой коры были исследования школы И. П. Павлова. Эти исследования показали, что разрушение коры затылочной области у собаки приводит к нарушению анализа и синтеза зрительных сигналов, в то время как разрушение коры височной области - к нарушению анализа и синтеза слуховых сигналов, и т. п. Таким образом, с помощью физиологич. исследований было показано, что отд. зоны коры являются высшими отделами соответствующих анализаторов: затылочная область является зрительной, теменная часть - общечувствительной, верхневисочная область - слуховой зоной коры головного мозга (см. рис. 3). Эти данные были подтверждены и наблюдениями над случаями разрушения (в результате ранения, кровоизлияния, опухоли) соответствующих отделов мозговой коры. Оказалось, что разрушение передней центральной извилины или ведущих к ней волокон приводит к параличу соответствующих конечностей противоположной стороны тела, разрушение задней центральной извилины - к нарушению чувствительности противоположной стороны тела, разрушение затылочной области - к выпадению или нарушению зрения, двустороннее поражение верхневисочной области - к выпадению или нарушению слуха.
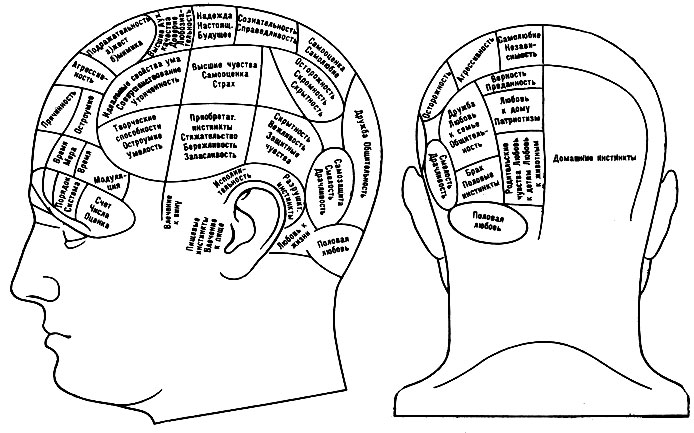
Рис. 1. Френологическая карта локализации психических способностей. Приводится по современной Ф. А. Галлю статуе
Положение о высокой специализации отд. участков мозговой коры было подтверждено и анатомич. данными. Ещё в 1874 киевский анатом В. А. Бец показал, что тонкое строение различных участков коры головного мозга является неодинаковым: кора передней центральной извилины имеет крупноклеточное строение, и в её толще заложены гигантские клетки пирамидной формы (т. н. пирамидные клетки Беца); в отличие от этого, кора задней центральной извилины имеет мелкоклеточное зернистое строение. Исследование тонкого анатомич. строения коры головного мозга было продолжено рядом выдающихся анатомов (К. Бродман, Экономо, О. Фохт, С. Рамон-и-Кахаль, Лоренте де Но, сотрудники Моск. ин-та мозга), работы к-рых позволили показать, насколько неоднородным является гистологич. строение зон мозговой коры. Как было показано дальнейшими исследованиями, эти отличия имеют существенное функциональное значение. Гигантские пирамидные клетки, расположенные в одном из нижних слоёв коры (в её 5-м слое), являются эфферентными (двигательными) клетками: их длинные отростки - аксоны - идут вниз, в продолговатый и спинной мозг и, переключаясь в передних рогах спинного мозга, направляются к мышцам. Возбуждение, возникшее в этих клетках, несёт двигательные импульсы, вызывая соответствующее сокращение мышц. Мелкоклеточные участки коры задне-центральной, затылочной и височной областей имеют др. функциональное значение. Мелкие зернистые клетки, расположенные в 4-м слое коры этих областей, являются приёмниками тех возбуждений, к-рые доходят до них от периферич. рецепторных приборов (тактильных, зрительных или слуховых). Волокна, начинающиеся в чувствительных приборах кожи, сетчатке глаза или в слуховых рецепторах внутреннего уха, идут в составе чувствительных нервов, проходят в подкорковые образования (специфич. ядра зрительного бугра и прилегающие к нему ядра коленчатых тел), а затем направляются к соответствующим отделам коры головного мозга (задней центральной извилине, полюсу затылочной области, внутренней поверхности верхней височной извилины); здесь эти афферентные волокна, несущие специфич. (осязательные, зрительные, слуховые) импульсы, кончаются в 4-м слое коры, а принесённые ими возбуждения передаются на расположенные в этом слое мелкие зернистые клетки.
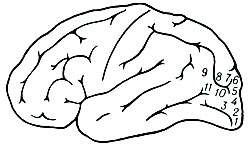
Рис. 2. Схема точек затылочной области, раздражение к-рых вызывает зрительные галлюцинации. Цифрами обозначены те участки мозговой коры, при раздражении которых возникали соответствующие зрительные галлюцинации: 1 - 'светящиеся шары'; 2 - 'окрашенный свет'; 3 - 'белый свет'; 4 - 'голубой диск'; 5 - 'пламя'; 6 - 'голубой туман'; 7 - 'лица, звуки, идущие со стороны вниз'; 8 - 'человек, идущий со стороны'; 9 - 'лица'; 10 - 'звери'; 11 - 'лица и бабочки'
Т. о., все указанные зоны коры головного мозга, непосредственно связанные с периферией длинными проекционными путями, являются высокоспецифическими проекционными (или первичными) зонами мозговой коры, одни из к-рых несут двигательную функцию, а другие - чувствительную.
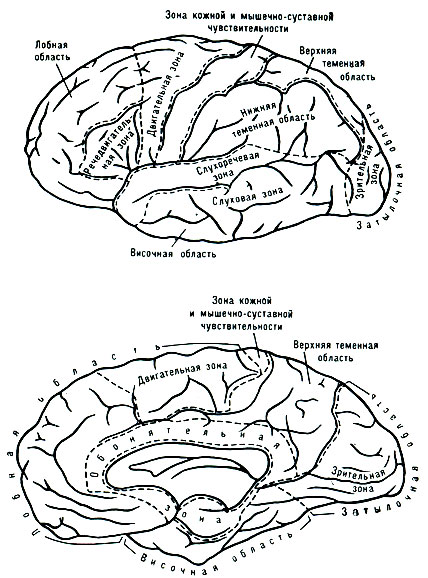
Рис. 3. Карта областей и полей коры головного мозга человека. Вверху - наружная поверхность полушарий; внизу - внутренняя поверхность полушарий
Характерно, что как тонкое строение проекционных зон мозговой коры, так и занимаемая каждой из них площадь отражают значение, к-рое имеет в жизни животного тот или иной периферич. аппарат. Так, двигательная область коры головного мозга животных с массивными и грубыми движениями отличается массивной величиной гигантских пирамидных клеток; наоборот, кора двигательной области животного с тонкими и сложными движениями отличается большей многочисленностью и меньшими размерами этих клеток. С другой стороны, чем большее значение имеет тот или иной периферич. чувствительный прибор или двигательный орган в жизни животного, тем большее место занимает та территория коры, в к-рой этот орган представлен. Так, в коре головного мозга ежа или крота, в жизни к-рых ведущее место занимает обоняние, обонятельная область коры занимает очень большое место, в то время как у человека, в жизни к-рого обоняние не играет столь существенной роли, обонятельная зона коры очень невелика. В коре головного мозга человека проекции руки, большого и указательного пальцев, мышц лица и языка занимают очень большое место, а проекции бедра, голени - небольшое место (см. рис. 4).
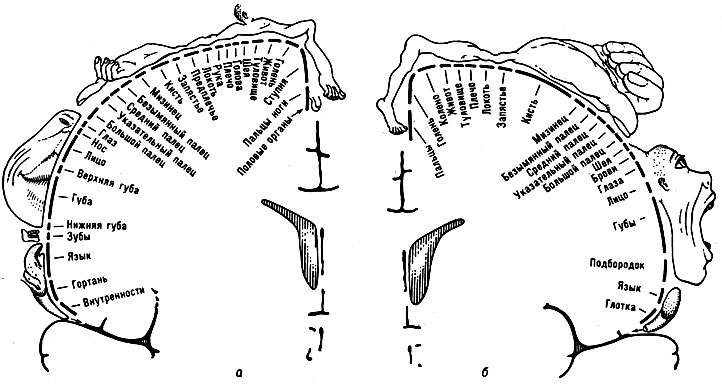
Рис. 4. а - корковая проекция чувствительности; б - корковая проекция двигательной системы. Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения
Проекционные (или первичные) зоны коры головного мозга человека являются аппаратами, лишь принимающими информацию (чувствительные проекционные зоны) или посылающими готовые импульсы на периферию (двигательные проекционные зоны). Переработка же принятой информации происходит при ближайшем участии более сложно построенных вторичных и третичных зон коры. Вторичные поля представляют собой области мозговой коры, непосредственно прилегающие к соответствующим проекционным (первичным) полям. Они отличаются тем, что волокна, приходящие к ним, не связывают их с периферией непосредственно, но несут импульсы, уже переработанные в особых ядрах зрительного бугра, а также тем, что в толще этих областей коры особенно мощное развитие получают верхние слои (2-й и 3-й), состоящие из нейронов с короткими отростками. Эти нейроны позволяют передавать возбуждения, дошедшие до воспринимающих клеток 4-го слоя, на соседние и отдалённые участки (причём такая передача происходит как по горизонтальным связям, расположенным внутри коры, так и по вертикальным связям, через соответствующие подкорковые образования). Эти нейроны, к-рые часто наз. ассоциативными, обеспечивают возникновение стойких и вместе с тем высокоподвижных кругов циркулирующего в коре возбуждения; они и являются физиологич. механизмами, к-рые обеспечивают многообразные контакты отд. нейронов и позволяют осуществлять сложнейшую аналитико-синтетич. работу мозговой коры и, по-видимому, являются физиологич. механизмами, лежащими в основе удержания следов тех возбуждений, к-рые поступают в мозг. Опыты, проведённые рядом исследователей во время мозговых операций, показывают, что раздражение проекционных зон зрительной или слуховой коры вызывает лишь появление элементарных ощущений (при раздражении проекционных зон зрительной коры - появление световых мельканий, пятен, при раздражении проекционных зон слуховой коры - появление тонов); раздражение же вторичных зон мозговой коры приводит к возникновению значительно более сложных психич. процессов.
Так, раздражение вторичных зон зрительной коры может вызвать оформленные предметные зрительные галлюцинации (см. выше, рис. 2), а раздражение вторичных зон слуховой коры - осмысленные слуховые галлюцинации (слышание музыки, речи и т. п.). Роль вторичных зон мозговой коры в осуществлении сложных психич. процессов подтверждается и наблюдениями над изменением психич. функций у больных с ограниченными поражениями этих отделов (ранение, кровоизлияние, опухоль). Так, если поражение проекционных зон зрительной коры приводит к частичному или полному выпадению зрения, то поражение вторичных зон вызывает нарушение синтетич. зрительного восприятия предметов и их узнавания (т. н. оптич. агнозия): хорошо воспринимая отд. признаки предметов, такой больной не может объединить их в единое целое и оказывается не в состоянии узнать знакомые изображения.
Т. о., первичные и вторичные зоны задних (теменных, затылочных и височных) отделов коры головного мозга обеспечивают работу по анализу и синтезу информации, пришедшей в кору головного мозга от осязательных, зрительных и слуховых воспринимающих приборов и представляют собой корковые отделы осязательного, зрительного и слухового анализаторов.
Познавательная деятельность человека не ограничивается, однако, работой изолированных анализаторов; уже каждое предметное восприятие предполагает синтез отдельных (осязательных, зрительных и слуховых) впечатлений или, иначе говоря, совместную работу этих анализаторов. Такая совместная работа осуществляется с помощью участия третичных зон коры головного мозга, или "зон перекрытия" корковых концов отдельных анализаторов. Эти зоны расположены на границе между теменными, затылочными, височными отделами коры головного мозга. Они либо ещё не существуют, либо относительно слабо развиты у высших млекопитающих и представляют собой специфически человеческие отделы коры головного мозга. В онтогенезе они развиваются значительно позже, чем первичные и вторичные зоны, и их развитие завершается у ребёнка лишь к 7 - 8-летнему возрасту. Для их строения характерно особенно мощное развитие верхних слоёв мозговой коры, содержащих наиболее сложные по своему строению ассоциативные нейроны. Клинич. наблюдения показывают, что поражение этих областей приводит к сложным изменениям синтетич. процессов, предполагающих совместную работу упомянутых анализаторов. Так, поражение теменно-затылочных отделов мозговой коры может привести к нарушению сложных форм ориентации в пространстве, конструктивной деятельности и к существенному затруднению выполнения всех интеллектуальных операций, к-рые протекают с участием пространственного анализа и синтеза (операций счёта, анализа сложных семантич. структур и т. п.).
Вторичные и третичные зоны коры головного мозга обоих полушарий неравноценны и играют существенно различную роль в осуществлении сложных форм познавательной деятельности. Одно из полушарий (левое полушарие у праворуких) связано с речевыми процессами и играет доминирующую роль в выполнении всех форм сложного (и особенно речевого) мышления; другое полушарие играет подчинённую роль, и его поражения не вызывают столь отчётливых нарушений. У левшей соотношение обоих полушарий носит обратный характер.
Роль третичных зон коры головного мозга в осуществлении сложных познавательных процессов ещё недостаточно изучена, и их изучение составляет предмет целого ряда нейропсихологич. исследований.
Описанный комплекс зон коры головного мозга, расположенный в пределах задних (теменно-височно-затылочных) отделов полушарий, обеспечивает анализ и синтез доходящей до мозга информации и является мозговой основой сложных познавательных операций. От него сильно отличается комплекс участков коры головного мозга, расположенный спереди от передней центральной извилины и входящий в состав центральных отделов двигательного анализатора. В отличие от задних отделов мозга, имеющих мелкоклеточное строение и входящих в систему анализаторов внешнего мира, в этих отделах коры преобладают большие и малые пирамидные клетки, и они характеризуются специфич. "вертикальной исчерченностью", указывающей на их ближайшее отношение к организации сложных двигательных процессов. Роль первичной зоны играет проекционная двигательная область коры. К ней непосредственно примыкает вторичная - премоторная зона коры, к-рая сохраняет строение, типичное для двигательной коры, но в к-рой преобладают "ассоциативные" клетки 2-го и 3-го слоев. Эти отделы называются "экстра- пирамидными двигательными полями". Их раздражение может привести к комплексным последовательным движениям, а их поражение ведёт к нарушению плавного перехода одного звена сложного движения в другое и к нарушению двигательных навыков. При массивных поражениях этой зоны возникают явления патологич. инертности двигательных процессов.
Спереди от премоторной зоны расположены поля лобной, или префронтальной, области мозговой коры. Их можно рассматривать как третичные поля, к-рые, входя в состав корковых отделов двигательного анализатора, вместе с тем сохраняют теснейшую связь с подкорковыми образованиями, а через них (и через посредство внутрикорковых длинных ассоциативных связей) и со всеми остальными отделами мозговой коры. Они относительно слабо развиты у животных и особенно мощно развиваются у человека, представляя собой специфически человеческие образования. Они также формируются в онтогенезе относительно поздно и завершают своё развитие лишь к началу школьного возраста. Несмотря на многочисленные исследования, функция лобных долей мозга остаётся недостаточно изученной. Раздражение этой области коры электрич. током не вызывает к.-л. отчётливых реакций; поражение этих областей патологич. процессом может не приводить к выпадению к.-л. видов чувствительности, не вызывает нарушений движения или речи. Однако массивное поражение лобных долей мозга может привести к отчётливым нарушениям сложных видов организованного поведения: больные перестают подчинять своё поведение возникающим намерениям, оказываются не в состоянии строить свои действия соответственно речевой инструкции и легко подпадают под влияние посторонних, отвлекающих их внимание раздражителей; сохраняя полученную ранее информацию, они перестают создавать сколько-нибудь сложные программы действий и регулировать действия соответственно этим программам, в связи с чем их деятельность легко теряет свой избирательный целенаправленный характер; они не могут сличать результат своего действия с исходным намерением и осознавать и корригировать допускаемые ошибки. Всё это заставляет предполагать, что лобные доли мозга имеют ближайшее отношение к функциям программирования, регуляции и контроля сложных форм психич. деятельности.
Особое место среди зон мозговой коры занимают те её отделы, к-рые расположены на внутренней и базальной поверхности больших полушарий (т. н. медиобазальные отделы мозговой коры, лимбич. область, гиппокамп и др.). Составляя часть наиболее древней мозговой коры, они, согласно совр. представлениям, не входят в состав тех отделов коры, к-рые обеспечивают получение информации или программирование движений и действий. Имея теснейшую связь с нижележащими образованиями мозгового ствола (т. н. ретикулярной формацией, гипоталамусом и др.), обеспечивающими общий тонус коры, они принимают ближайшее участие в регуляции состояний коры. Поэтому патологич. состояния этих отделов мозга, не отражаясь на познавательных процессах или двигательных актах, могут вызвать заметные изменения общего активного состояния субъекта. Поражение медиальных отделов височной области (гиппокампа) может вызвать массивные изменения памяти и привести к снижению бодрственного состояния. Поражение базальных отделов лобной области может привести к заметному нарушению в регуляции влечений и к общим изменениям активности субъекта. Функция медиобазальных отделов коры головного мозга стала предметом тщательного изучения лишь в последнее время, и детальный анализ её роли в протекании психич. деятельности человека ещё отсутствует.
Описанные представления о структурной и функциональной организации коры головного мозга человека позволяют на новых науч. основах подойти к вопросу о Л. п. ф. в коре головного мозга человека. В отличие от старых, не оправдавших себя представлений, согласно к-рым психич. процессы рассматривались как первичные и далее неразложимые свойства психики ("способности"), совр. науч. психология подходит к психич. процессам как к сложившимся в социально-историч. развитии функциональным системам, к-рые имеют рефлекторное строение, сложный психофизиологич. состав и опираются на динамич. комплексы совместно работающих зон коры головного мозга, каждая из к-рых участвует в осуществлении этой сложной функциональной системы на своих собственных ролях. Именно поэтому сложные психич. функции не могут быть "локализованы" в изолированных участках коры головного мозга, но осуществляются целой системой зон, иногда далеко расположенных друг от друга. Так, осуществление сложного произвольного движения требует участия заднетеменных отделов коры (обеспечивающих кинестезич. основу двигательного акта), теменно-затылочных (позволяющих сохранять чёткие пространственные координаты движения), премоторных (дающих возможность осуществить сложные кинетич. мелодии с плавным переключением одного двигательного акта на другой), лобных отделов мозга (обеспечивающих осуществление сложных двигательных программ, постоянную регуляцию движений и контроль над их результатами). Вместе с тем всякое сложное произвольное движение может быть выполнено лишь при сохранении нормальных соотношений между корой и подкорковыми образованиями, влияние к-рых обеспечивает нужный тонус и координированность двигательных актов.
Ещё более сложными являются мозговые механизмы высших психич. функций, протекающих с участием речи и опирающихся на вторую сигнальную систему действительности. Такие процессы, как письмо, чтение, логич. запоминание и речевое мышление, осуществляются всегда сложной системой совместно работающих зон мозговой коры. Так, процесс письма осуществляется при ближайшем участии вторичных отделов коры левой височной области (обеспечивающих слуховой анализ и синтез речевых звуков), нижних отделов коры левой теменной области (создающих кинестезич. основу для артикуляции звуков речи, участвующих в письме), коры левой затылочно-теменной области (позволяющей перешифровывать звуки в графемы и сохранять пространственную организацию записываемых букв), нижних отделов левой премоторной области (участвующих в переключении с одного элемента записываемого слова на другой и позволяющих осуществлять запись слова как единой последовательной системы звеньев), лобной области (контролирующей общую программу процесса письма и обеспечивающей торможение побочных ассоциаций и контроль нужного действия) и т. д. Совершенно естественно поэтому, что нарушения процесса письма могут возникнуть при поражении самых различных по локализации участков коры левого полушария и каждый раз нарушения письма будут носить различный характер. Естественно, вместе с тем, что письмо, имеющее различное психологич. строение (напр., звуко-буквенное письмо на рус. яз. и иероглифич. письмо на др.-егип. или кит. яз.), будет осуществляться на основе работы различных зон коры головного мозга: слуховые отделы коры, необходимые для выделения, анализа и синтеза звуков при письме на рус. яз. будут не столь необходимы для иероглифич. записи понятий (минующих анализ звукового состава слова) на кит. яз. Следует также отметить, что письмо начинающего ученика (когда развёрнутый звуковой анализ играет ведущую роль), по-видимому, опирается на иное соотношение мозговых зон, чем автоматизированное письмо, при к-ром ведущую роль могут играть хорошо упроченные кинестезич. стереотипы.
Значение тех или иных зон мозговой коры для осуществления и развития психич. функций не остаётся одинаковым на различных этапах возрастного развития, и поражение одних и тех же зон мозговой коры у взрослого человека и на ранних этапах психич. развития ребёнка может иметь различные результаты. Так, поражение вторичных отделов затылочных областей коры у взрослого приводит к ограниченным нарушениям сложного зрительного восприятия; поражение тех же отделов в раннем возрасте может привести к иным результатам: нарушая процессы наглядного анализа и синтеза, оно нарушает основу, на к-рой складывается дальнейшее интеллектуальное развитие ребёнка, и может вызвать в качестве вторичного эффекта тяжёлое недоразвитие интеллектуальных процессов.
Положение о том, что оценка роли, к-рую играет та или иная зона мозговой коры в психич. деятельности, возможна только при учёте знания осн. законов психич. развития, имеет решающее значение для науч. представления о Л. п. ф. в коре головного мозга.
Лит.: Бехтерев В. М., Основы учения о функциях мозга, СПБ, 1903 - 07, в. 1 - 7; Павлов И. П., Лекции о работе больших полушарий головного мозга. Полн. собр. соч., 2 изд., т. 4, М., 1951; Пенфилд [У.] и Эриксон Т., Эпилепсия и мозговая локализация, пер. с англ., [М.], 1949; Пенфилд У. и Джаспер Г., Эпилепсия и функциональная анатомия головного мозга, М., 1958; Выготский Л. С., Психология и учение о локализации психических функций, в его кн.: Развитие высших психических функций, М., 1960; Лурия А. Р., Высшие корковые функции человека и их нарушения при локальных поражениях мозга, М., 1962.
Источники:
- Педагогическая энциклопедия/Глав. ред. И. А. Каиров и Ф. Н. Петров. т. 2. - М.: Советская энциклопедия, 1965. - 912 с. с илл., 5 л. илл.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://pedagogic.ru/ 'Библиотека по педагогике'